

Одним из характерных свойств сердца является его автоматия. Автоматией называется способность органа, ткани или клетки возбуждаться под влиянием импульсов, возникающих в них самих без внешних раздражителен. Причиной, вызывающей автоматию, являются изменения обмена веществ, происходящие в самом органе или клетке.
Изолированное сердце. Автоматию сердца проще всего наблюдать в опытах на изолированном, т. е. вырезанном из организма сердце лягушки. Такое изолированное сердце лягушки, помещенное в раствор Рингера, может сокращаться много часов и даже дней.
Для того чтобы сокращалось изолированное сердце теплокровного, применяется методика Лангендорфа: в аорту вырезанного сердца вставляют канюлю, соединенную с высоко расположенным стеклянным сосудом, наполненным дефибринированной кровью или раствором Рингера, к которому добавлена глюкоза; раствор Рингера насыщают кислородом и нагревают до 37—38°. Под давлением столба жидкости, притекающей в аорту из сосуда с раствором Рингера, полулунные клапаны аорты закрываются, и раствор течет в питающие сердце венечные артерии. В этих условиях сердце теплокровного может часами ритмически работать.
Применяя методику нропускапия раствора через сосуды органа (методика перфузии), можно восстановить сокращения уже остановившегося сердца спустя несколько часов после смерти животного или человека. Впервые опыты с оживлением сердца ребенка были произведены А.А. Кулябко в 1902 г. В дальнейшем последовали успешные опыты по восстановлению работы вырезанного из трупа сердца взрослого человека почти через 2 суток после смерти. Оживленное сердце билось свыше 13 часов (С. В. Андреев).
Автоматия разных отделов сердца. Наибольшей способностью к автоматии обладает водитель ритма нормального сердца — синоатриальный узел. Однако способность к автоматии присуща и некоторым другим участкам сердца. Так, автоматия обнаружена в волокнах близких по структуре к волокнам синоатриального узла, расположенных вблизи него в предсердиях. Эти волокна обладают несколько меньшей способностью к автоматии, чем клетка синоатриального узла. Их называют скрытыми водителями ритма так как в норме их автоматия не проявляется и они берут на себя эту функцию лишь после того, как нарушается функция основного водителя ритма.
Автоматия свойственна также атриовентрикулярному узлу. В этом можно убедиться путем экспериментов на собаке с наложением лигатуры выше атриовенгрикулярного узла. В этом случае отключаются предсердные водители ритма и желудочек сначала перестает сокращаться, но по прошествии некоторого времени сокращения его возобновляются за счет автоматии атриовенгрикулярного узла. То же самое наблюдается при сильном охлаждении области синоатриального узла: прекратившиеся сокращения желудочков восстанавливаются под влиянием импульсов, возникающих в атриовентрикулярном узле. При этом сокращения предсердий и желудочков могут происходить не в обычной последовательности, одно за другим, а почти одновременно, потому что возбуждение из атриовентрикулярного узла одинаково быстро достигает мышцы предсердия и желудочка (атриовентрикулярный ритм сокращений сердца).
После отделения лигатурой атриовентрикулярного узла от ниже расположенных участков проводящей системы все же происходят сокращения желудочков. В таком случае водителями ритма становятся обладающие автоматной волокна Пуркине в правом или левом желудочке.
Синоатриальный узел называют центром автоматии первого порядка, атриовентрикулярный узел — центром автоматии второго порядка. Частота сердечных сокращений, задаваемая синоатриальным водителем ритма, у человека в покое в среднем равна 70 —75 в минуту. При атриовентрикулярном ритме она примерно вдвое ниже, а в том случае, когда сердце работает под влиянием автоматии ниже расположенных водителей ритма, частота сокращений желудочков еще меньше.
Частота автоматически возникающих возбуждений рассматривается как показатель степени автоматии водителя ритма. Упомянутые выше различия в частоте возбуждений сердца, генерируемых разными водителями ритма, указывают на то, что самой высокой автоматией обладает синоатриальный узел, меньшей — атриовентрикулярный узел и еще меньшей — волокна Пуркине, находящиеся в желудочках сердца. Таким образом, чем дальше расположен очаг автоматии от венозного конца сердца и чем ближе он находится к артериальному концу, тем меньше его способность к автоматии. Эта зависимость получила название убывающего градиента автоматии (Гаскелл).
В нормальных физиологических условиях функционирует лишь один очаг автоматии — синоатриальный узел. Водители ритма второго и третьего порядка при этом «безмолвствуют», т. е. их способность к автоматии не проявляется. Это объясняется тем, что их автоматия подавлена приходящим к ним от выше расположенного водителя ритма более частым ритмом импульсов, чем тот, который они сами способны генерировать. После выключения синоатриального узла (наложением лигатуры, охлаждением, введением некоторых ядов), когда ритмический поток импульсов к атриовентрикулярному узлу и волокнам Пуркине прекращается, восстанавливается автоматия этих водителей ритма. На восстановление их автоматии требуется некоторое время, которое получило название преавтоматической паузы. Длительность последней равна от нескольких секунд до нескольких десятков секунд, в течение которых наблюдается асистолия, т. о. прекращение сокращений сердца.
Угнетение автоматии желудочковых водителей ритма частым ритмом возбуждений можно видеть в следующем эксперименте: если после наложения лигатуры на область атривентрикулярного узла на фоне редкого ритма сокращении желудочков раздражать их ритмическими электрическими стимулами большой частоты, то желудочки воспроизводят ритм раздражений. Однако их автоматия подавляется и при выключении электрической стимуляции желудочки перестают сокращаться в течение некоторого времени, пока восстановится их собственная автоматия.
Анатомический субстрат и природа автоматии. У взрослых животных автоматия свойственна волокнам атипической мускулатуры, сосредоточенным в проводящей системе сердца.
Ритмическое возникновение импульсов без каких-либо ритмических раздражений извне можно наблюдать в клетке сердца, отделенной от других. Это доказано при культивировании вне организма отдельных клеток сердца. Для этой цели вырезают кусочек сердца молодого животного, например крысенка, и подвергают его на короткое время действию пищеварительного сока, который переваривает в первую очередь внеклеточные белки, являющиеся как бы цементом, соединяющим отдельные клетки, но не успевает разрушить сами клетки. Отделенные таким способом друг от друга клетки промывают сывороткой крови и помещают в стеклянной чашечке в термостат, где они находятся при температуре 37°. Сыворотку крови, служащую питательным материалом, периодически заменяют.
Через несколько часов некоторые клетки, примерно одна из 100, начинают ритмически сокращаться с частотой от 10 до 150 сокращений в минуту. Автоматик) таких культивируемых вне организма клеток удавалось поддерживать до 40 дней. За это время отдельные клетки соединялись друг с другом, образуя сети, росли и делились. Если в культуре имеется несколько сокращающихся клеток, то их сокращения могут происходить в разном ритме. Однако если между клетками устанавливается анатомическая связь, то они начинают сокращаться в одном и том же ритме, свойственном той клетке, которая сокращалась чаще. Очевидно, эта клетка, обладая более высокой автоматией, подавляет способность к автоматии других клеток. При разделении группы синхронно сокращающихся клеток на две половины каждая из них начинает сокращаться с разной частотой.
Описанные опыты доказывают, что анатомическим субстратом автоматии являются некоторые мышечные клетки сердца; это находится в соответствии с так называемой миогенной теорией автоматии сердца.
Электрофизиологические исследования, проведенные с помощью внутриклеточного микроэлектрода, обнаружили характерную особенность мышечных волокон-водителей ритма сердца. Эта особенность состоит в том, что в промежутке между двумя сокращениями в диастолу в автоматически возбуждающейся клетке происходит постепенное уменьшение мембранного потенциала, т. е. разности потенциалов между протоплазмой и внешней поверхностью клетки. Когда разность потенциалов уменьшается до определенного критического уровня, внезапно возникает крутой сдвиг электрического заряда клетки, что свидетельствует о ее возбуждении. Возникшая волна возбуждения распространяется на другие, сначала близлежащие, а затем и отдаленные клетки, которые также приходят в состояние возбуждения.
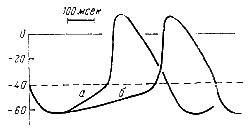 |
Благодаря этому автоматически возбуждающаяся клетка становится водителем ритма сердца. Для того чтобы возникло распространяющееся возбуждение, мембранный потенциал должен уменьшиться на 20—30 мв. Чем быстрее достигается во время диастолы такое изменение мембранного потенциала, тем чаще возникает возбуждение клетки-водителя ритма и тем, следовательно, чаще будет происходить сокращение сердца (рис. 11). Из этих фактов следует, что автоматия клеток-водителей ритма связана с изменениями их электрического состояния. Рис. 11. Схема, показывающая зависимость частоты возбуждении сердца от крутизны изменения мембранного потенциала клетки-водителя ритма во время диастолы (по Гоффману и Кренфильду). Пунктиром показан критический уровень мембранного потенциала. |
Автоматическое изменение внутриклеточного электрического потенциала во время диастолы характерно для клеток синоатриального узла, причем обычно одновременно возбуждается несколько клеток, лежащих в разных частях узла. Это означает, что автоматия узла имеет многоочаговый характер. В клетках сердца, в данное время не выполняющих функции водителя ритма, например в волокнах атрновентрикулярного узла и в волокнах Пуркине, не отмечается спонтанного медленного изменения потенциала во время диастолы. Однако если к этим клеткам перестают поступать импульсы из синоатриального узла, то в них начинает происходить спонтанное уменьшение разности потенциалов до такого уровня, по достижении которого возникает возбуждение. Таким образом, и этих клетках пробуждается автоматия, и они становятся водителями ритма сердца.
Пока совершенно неясно, чем вызвано ритмическое, спонтанно возникающее изменение заряда клетки, лежащее в основе автоматии сердца. Имеются указания, что определенная роль в автоматии принадлежит ацетилхолину, содержание которого в клетках-водителях ритма выше, чем в других мышечных волокнах сердца. В клетках синоатриального и атриовентрикулярного узлов содержание натрия больше, чем в сократительном миокарде, что может быть важное в связи с тем большим значевнием, которое имеют эти ионы в процессе возбуждения. По-видимому, автоматия обусловлена особенностями процессов обмена веществ в клетках-водителях ритма, изменяющими состояние их поверхностных мембран.
