

Возникающий в возбужденном участке потенциал действия становится раздражителем для соседних покоящихся участков нервного или мышечного волокна. Измерения показывают, что в различных волокнах величина потенциала действия (около 110—120 мв) в 5—10 раз превышает порог раздражения. Это обеспечивает бездекрементное (т. е. незатухающее) распространение возбуждения вдоль возбудимой мембраны.
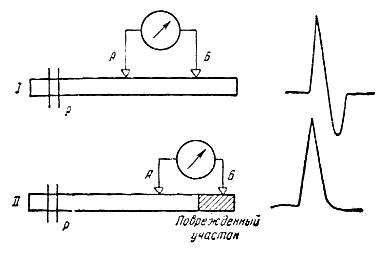 |
Для того чтобы зарегистрировать распространие возбуждения в нервном волокне, необходимо к двум его точкам А и Б приложить электроды, соединенные с осциллографом, а точке Р нанести достаточно сильный раздражающий стимул. При таком отведении на экране осциллографа можно получить двухфазный потенциал действия, подобный тому, который приведен на рис. 131, I. Рис. 131. Схема расположения раздражающих (Р) и отводящих (А и Б) электродов для отведения двухфазного (I) и монофазного (II) потенциалов действия. |
Схема на рис. 132 разъясняет механизм возникновения этих двух противоположно направленных колебаний потенциала. В состоянии покоя все точки наружной поверхности возбудимой мембраны заряжены электродположительно по отношению к ее внутренней поверхности. Когда волна возбуждения проходит по участку под электродом, ближайшим к месту раздражения, наружная поверхность мембраны в этом участке становится электроотрицательной по отношению к точке Б. Это вызывает отклонение луча осциллографа вверх. Когда волна возбуждения покидает этот участок, луч возвращается в исходное положение.
Затем возбуждение достигает участка под вторым электродом Б; этот участок в свою очередь становится электроотрицательным по отношению к точке А и луч осциллографа отклоняется вниз. Распространение возбуждения становится причиной возникновения двухфазности потенциала действия. Если же умертвить участок нерва под дальним электродом Б какими-либо агентами, например холодом или новокаином, или нарушить проведение возбуждения между участками А и Б, то вторая фаза потенциала действия не возникает, регистрируемый потенциал действия будет однофазным (рис. 131, II)
Механизм проведения возбуждения от одного участка волокна к другому в принципе не отличается от механизма возникновения импульса в точке раздражения. В обоих случаях потенциал действия возникает, когда деполяризация мембраны достигает критической величины. Однако если при искусственном раздражении эта деполяризация осуществляется катодом извне приложенного тока, при проведении возбуждения деполяризацию мембраны вызывает электрический ток, возникающий между возбужденным (электроотрицательным) и покоящимся (электроположительным) участками мембраны.
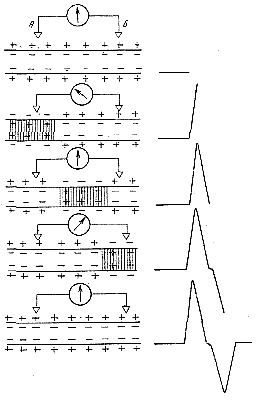 |
Это может быть доказано следующим опытом. Если участок нервного волокна между раздражающими и отводящими электродами поместить в жидкую среду, лишенную ионов и, следовательно, обладающую очень высоким сопротивлением для электрического тока (такой средой может быть, например, изотонический раствор сахарозы), то проведение волны возбуждения через этот участок резко замедляется или даже полностью прекращается. Скорость распространения возбуждения варьирует в различных нервных и мышечных волокнах в широких пределах. В нервных волокнах теплокровных животных она достигает величин порядка 120 м/сек; в нервных волокнах моллюска анодонты скорость проведения составляет лишь 0,01 м/сек. Рис. 132. Схема, иллюстрирующая механизм возникновения двухфазного потенциала действия |
