

Н. Е. Введенский в 1896 г. наблюдал, что раздражение участка двигательной зоны коры больших полушарий головного мозга, вызывающего сокращение мышц-сгибателей противоположной передней конечности, приводит к расслаблению мышц-разгибателей этой конечностри одновременно к сокращению мышц-разгибателей передней конечности на стороне раздражения. Эти явления объясняются тем, что при возбуждении центра сгибательной мускулатуры одной конечности происходит торможение центра разгибательной мускулатуры той же конечности и возбуждение центра мышц-разгибателей второй конечности.
Ч. Шеррингтоном установлены аналогичные взаимоотношения между центрами мышц-сгибателей и мышц-разгибателей у спинального животного, у которого спинной мозг отделен от головного. Так, если кожу лапы спинальной кошки раздражать электрическим током, то это вызовет сгибательный рефлекс: сокращение мышц-сгибателей и расслабление мышц-сгибателей лапы. Одновременно происходит перекрестный разгибательный рефлекс, состоящий в сокращении мышц-разгибателей и расслаблении мышц-сгибателей лапы противоположной стороны. Эти явления объясняются тем, что при возбуждении центра, вызывающего сгибание одной конечности, происходит торможение центра мышц-разгибателей этой конечности. Одновременно возбуждается центр мышц-разгибателей и тормозится центр мышц-сгибателей второй симметричной конечности.
Таким образом, центры мышц-антагонистов — сгибателей и разгибателей — находятся при выполнении многих двигательных актов в противоположном состоянии. Лишь при этом возможно точное движение сгибания или разгибания. Анализ подобных явлений привел к представлению о реципрокной, или сопряженной, иннервации мышц-антагонистов. Согласно этому представлению, возбуждение центра одной группы мышц сопровождается реципрокным торможением (сопряженным) центров антагонистической мускульной группы.
Механизм реципрокного торможения спинномозговых двигательных центров был выяснен в сравнительно недавнее время исследованиями Экклса и его сотрудников (рис. 184). Афферентные нервные волокна— длинные отростки биполярных клеток спинального ганглия — дают в спинном мозгу разветвления, одни из которых возбуждают мотонейроны, иннервирующие мышцы-сгибатели, а другие — нейроны, образующие тормозные синапсы на мотонейронах, иннервирующих мышцы-разгибатели. Таким образом, раздражение афферентного волокна вызывает одновременно возбуждение центра мышц-сгибателей и торможения центра мышц-разгибателей.
Предполагают, что на сходном механизме основано сопряженное торможение и других, более сложных рефлексов: афферентный стимул, возбуждающий данный рефлекторный центр через посредство тормозных промежуточных нейронов, угнетает деятельность других центров. Например, при осуществлении того или иного сложного рефлекторного акта (пищевого, полового, оборонительного и др.) у животных и человека другие рефлексы, как правило, ослабляются или даже полностью затормаживаются. Так, сильное болевое раздражение, нанесенное во время акта еды, вызывает оборонительную реакцию и тормозит пищевой центр. Напротив, сильное возбуждение пищевого центра у голодного животного приводит к понижению возбудимости центров оборонительного рефлекса, вследствие чего животное становится менее чувствительным к слабым болевым раздражениям.
Взаимотормозящие влияния рефлексов, основанные на реципрокном торможении, называют часто одновременной отрицательной индукцией.
Следует подчеркнуть, что реципрокные отношения между различными центрами не являются абсолютно постоянными. Так, описанные выше взаимоотношения спинномозговых центров сгибания и разгибания в наиболее четкой форме могут быть выявлены только у спинального животного. В нормальных же физиологических условиях в целостном организме, как показал И. С. Беритов, реципрокные отношения между спинномозговыми центрами могут изменяться и подавляться под влиянием лабиринтных и шейных проприоцептивных рефлексов, центры которых находятся в стволовой части головного мозга. Координация этих центров в свою очередь подчинена влияниям, исходящим из подкорковых образований и коры больших полушарий головного мозга. Вследствие этого возможна большая изменчивость реакции, а поэтому у нормального животного возможны движения с точки зрения принципа реципрокной иннервации несовместимые, например одновременное сгибание и разгибание симметричных конечностей.
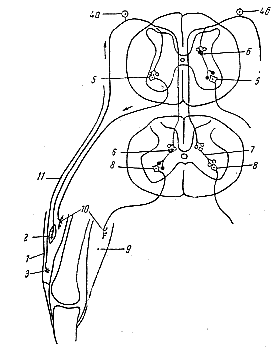 |
Рис. 184. Схема, иллюстрирующая механизм реципрокного торможения при рефлексах, вызванных раздражением мышечного веретена (слева) или тельца Гольджи в сухожилиях (справа) (по Дж. Экклсу). 1 - четырехглавая мышца бедра; 2 - мышечное веретено; 3 - тельце Гольджи в сухожилии; 4 - реценторные клетки в спинномозговом ганглии (4а — нервная клетка, воспринимающая импульсы от веретена; 4б - нервная клетка, воспринимающая импульсы от тельца Гольджи); 5 - мотонейроны, иннервирующие мышцы-разгибатели; 6 - тормозной промежуточный нейрон; 7 -возбуждающий промежуточный нейрон; 8 - мотонейроны, иннервирующие мышцы-сгибатели; 9 - мышца-сгибатель; 10 - моторные нервные окончания в мышцах; 11 - нервное волокно от тельца Гольджи. |
