

В течение многих лет шли споры о том, существует ли протоплазматическая непрерывность между нервными клетками в центральной нервной системе или же они структурно отделены друг от друга. В настоящее время установлено, что протоплазматическая непрерывность между отростками нервных клеток существует только у низших беспозвоночных животных — кишечнополостных, у которых нервная система построена по типу синцития. Такой синтиций получил название нейропиля. Нейропиль проводит возбуждение диффузно, причем оно может распространяться любом направлении.
У высших беспозвоночных животных и у всех позвоночных нервные клетки в центральной нервной системе связаны другс другом только синапсами.
 |
Аксон каждого нейрона, подходя к другим нервным клеткам, ветвится и образует многочисленные окончания на телах этих клеток и их дендритах. Так, на теле и дендритах крупной моторной нервной клетки передних рогов спинного мозга или пирамидной клетки коры полушарий головного мозга имеются тысячи нервных окончаний — синапсов, образованных нервными отростками многих других нейронов (рис. 166). Одно нервное волокно может образовать до 10 000 синапсов на многих ревных клетках. Синапсы, расположенные на теле (соме) нейрона, называют аксо-соматическими,а на дендритах — аксодендритными. Синапсы в центральной нервной системе, так же как и периферические синапсы, состоят из собственно нервного окончания, синаптической щели и постсинаптической мембраны. Рис. 166. Расположение синапсов на теле нейрона и его дендритах (по Хаачу и Барру). |
Нервные окончания в центральной нервной системе имеют вид пуговок, колечек или бляшек. Как показывают электронные микрофотографии (рис.167), каждая синаптическая пуговка или бляшка покрыта мембраной-так называемой пресинаптической мембарной, а внутри пуговки или бляшки находятся пузырьки, содержащие, согласно распространенным представлениям, возбуждающий или тормозной медиатор.
 |
Пресинаптическая мембрана отделена синаптической щелью от постсинаптической мембраны, находящейся на теле или дендритах нейрона, к которому передается нервный импульс. В зависимости от характера производимого эффекта синапсы подразделяют на возбуждающие и тормозные. Ниже мы рассмотрим принципиальный механизм передачи возбуждения через возбуждающие синапсы, механизм же действия тормозных синапсов опишем в разделе, посвященном центральному торможению. Рис. 167. Небольшой участок поверхности тела нервной клетки с синаптическими бляшками (А) и продольный разрез через одну синаптическую бляшку (В). Показаны синаптические пузырьки и митохондрии (по Уайкову и Юнгу). |
Механизм передачи возбуждения в синапсах центральной нервной системы. Медиатор, вырабатывающийся в нервных окончаниях этих синапсов, под влиянием поступающего к ним нервного импульса освобождается из связанного состояния и выбрасывается в синаптическую щель. Поскольку ширина этой щели очень невелика (около 200 Å), медиатор быстро (примерно в течение 0,5 мсек) диффундирует к постсинаптической мембране и взаимодействует с ее структурными компонентами — определенными белково-липоидными комплексами. В результате происходит значительное повышение проницаемости постсинаптической мембраны для ионов натрия и калия.
 |
Это приводит к деполяризации постсинаптической мембраны и появлению так называемого возбудительного постсинаптического потенциала. В момент, когда этот потенциал достигает критического уровня, в клетке возникает распространяющееся возбуждение — потенциал действия, или пиковый потенциал (рис. 168). Так как мембрана начального сегмента аксона обладает наиболее низким критическим уровнем деполяризации, то потенциал действия прежде всего возникает в этом участке нейрона и уже отсюда распространяется как на тело клетки, так и вдоль аксона. Рис. 168. Возникновение потенциала действия при достижении возбуждающим постсинаптическим потенциалом критической величины (по Дж. Экллсу). 1-3 - возбуждающие постсинаптические потенциалы при подпороговой силе афферентного раздражения; 4 — возникновение потенциалов действия при пороговой силе раздражения. Критический уровень деполяризации показан стрелкой. |
Если возбудительный постсинаптический потенциал, вызываемый афферентным раздражением, имеет большую продолжительность, в нервной клетке может возникнуть ритмический разряд следующих друг за другом потенциалов действия (рис. 169, А).
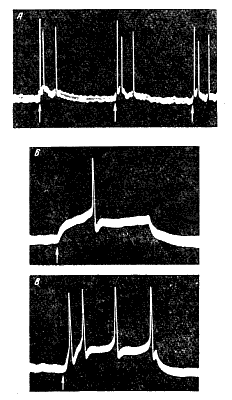 |
Многие клетки в центральной нервной системе обладают атвоматией, генерируют ритмические импульсы даже в отсутствие внешних раздражений. С помощью внутриклеточных микроэлектродов в таких клетках можно обнаружить постоянные медленные колебания мембранного потенциала, сопровождающиеся одиночными множественными разрядами потенциалов действия. Эти разряды возникают, когда мембрана деполяризуется до определенного критического уровня. Поступление нервных импульсов возбуждающим синапсам к такой автоматически разряжающейся клетке водит к усилению деполяризации мембраны и как следствие к увеличению частоты разрядов нервных импульсов. Природа возбуждающих медиаторов центральной нервной системе выяснена пока еще недостаточно. Только в отношении некоторых возбуждающих синапсов в головном и спинном мозгу установлено, что медиатором в них является ацетилхолин. Известно также, что при сильном возбуждении в ткани головного и спинного мозга увеличивается содержанне некоторых других высоко активных соединений — норадреналина, серотонина, вещества Р, которые, возможно, также являются медиаторами. Наличие в центральной нервной системе синапсов и химической передачи импульсов в них обусловливает ряд особенностей проведения возбуждения. Рис. 169. Ответы мотонейрона на одиночное раздражение афферентного нерва (А) и на непосредственное раздражение мотонейрона постоянным током (Б и В) (по А. И. Шаповалову). Микроэлектродное отведение. Б — ответ при пороговой силе раздражающего тока; В-ответы при сверхпороговой силе тока. Моменты стимуляции отмечены стрелками. |
