

Явление центрального торможения было открыто И. М. Сеченовым в 1862 г. Основной его опыт состоял в следующем. У лягушки делали разрез головного мозга на уровне зрительных бугров и удаляли большие полушария. После этого измеряли время рефлекса отдергивания задних лапок при погружении их в раствор серной кисл (методика Тюрка).
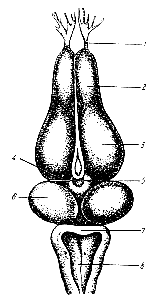 |
Этот рефлекс осуществляется спинномозговыми центрами и его время является показателем возбудимости центров. И. М. Сеченов обнаружил, что если на разрез зрительных бугров (рис. 177) наложить кристаллик поваренной соли или нанести слабое электрическое раздражение на эту область мозга, то время рефлекса резко удлиняется. На основании этого факта И. М. Сеченов пришел к заключению, что в таламической области мозга у лягушки существуют нервные центры, оказывающие тормозящие влияния на спинно-мозговые рефлексы. И. М. Сеченов правильно оценил важное значение открытого им явления центрального торможения и в своих теоретических работах исполь зовал его для объяснения физиологических механизмов поведения человека. Рис. 177. Головной мозг лягушки и линия разреза его в опыте И. М. Сеченова. 1 — обонятельный нерв; 2 — обонятельная доля; 3 — большие полушария; 4 — зрительный бугор; 5 — линия разреза головного мозга; 6 — двухолмие; 7 — мозжечок; 8 — продолговатый мозг и ромбовидная ямка. |
Вскоре были открыты новые факты, демонстрирующис явления торможения в центральной нервной системе. Ф. Гольц показал, что у лягушки рефлекс отдергивания задней лапки в ответ погружение ее в раствор кислоты может быть заторможен одновременным сильным механическим раздражением второй лапки, например сжатием ее пинцетом. Ф. Гольц установил также, что квакательный рефлекс лягушки, наблюдаемый надавливании на боковые стенки туловища, тормозится раздражением лапок.
Ф. Гольц наблюдал торможение спинномозговых рефлексов и после удаления у лягушек таламической области, и поэтому он выступил против представления о существовании в стволе мозга особых тормозящих центров. Гольц считал, что торможение может развиться в любом отделе нейтральной нервной системы при встрече двух или нескольких раздражений, вызывающих различные рефлексы.
Ч. Шеррингтоном, Н. Е. Введенским, А. А. Ухтомским и многими другими исследователями было показано, что торможение играет важную роль в деятельности всех отделов центральной нервной системы.
Приведем некоторые примеры внутрицентрального торможения из работ Ч. Шеррингтона, обстоятельно изучившего закономерности взаимодействия процессов возбуждения и торможения в спинном мозгу млекопитающих животных.
У кошки с удаленными большими полушариями раздражают центральный копец n. popliteus, что вызывает рефлекторное сокращение мышцы разгибателя колена — m. vastus сrureus — противоположной конечности (рис. 178). Данный рефлекс имеет длительное последействие. Если во время него нанести второе раздражение на тот же n. popliteus, то возникает торможение ранее вызванного рефлекса и расслабление мышцы.
Рис. 178. Рефлекторное торможение сокращения m. vastus cruris при
повторном раздражении n. popliteus (слева) и при сильном раздражении кожи
одноименной стороны (справа) (по Ч. Шеррингтону). На левом рисунке: 1 —
отметка времени; 2— сокращение мышцы; 3 — отметка раздражения n.
popliteus, вызвавшего рефлекторное сокращение мышцы (А) и рефлекторное
торможение (Б). На правом рисунке: 1 — запись сокращения мышцы; 2— отметка
раздражения кожи на лапе одноименной стороны; 3 — отметка раздражения кожи
на лапе противоположной стороны; 4 — отметка времени (1/5
секунды).

Рефлекторное сокращение мышцы разгибателя колена можно вызвать раздражением кожи лапы противоположной стороны (перекрестный разгибательный рефлекс). Нанесение сильного раздражения на кожу лапы одноименной стороны сопровождается резким рефлекторным расслаблением этой мышцы вследствие возникшего в центрах торможения (рис. 178). Равным образом сгибательныи рефлекс у кошки, вызванный раздражением нерва одноименной стороны, тормозится раздражением нерва или кожи симметричной стороны.
Интенсивность рефлекторного торможения зависит от соотношения силы раздражений — возбуждающего и тормозящего нервный центр.
Если раздражение, вызывающее рефлекс, сильное, а тормозящее раздражение слабое, то интенсивность торможения невелика. При противоположном соотношении силы этих раздражении рефлекс будет полностью заторможен.
Если на нерв наносится несколько слабых тормозящих нервный центр раздражений,
то торможение оказывается усиленным, т. е. наблюдается щммация тормозных
влияний.
Н. Е. Введенский наблюдал явления торможения в коре больших
полушарий головного мозга. В его опытах на фоне раздражения определённой точки
двигательной зоны коры одного полушария (это вызывало сгибание одной из лап
противоположной стороны тела) раздражалась симетричная точка коры другого
полушария. В результате эффект первого
раздражения тормозился (согнутая лапа разгибалась).
Крупнейший вклад в учение о центральном торможении внес И. П. Павлов, изучивший торможение условных рефлексов и показавший, что явления торможения имеют важное значение во всех проявлениях высшей нервной деятельности и поведения организма.
По вопросу о механизме центрального торможения высказывались различные, внешне противоречивые представления. Одни исследователи полагали, что в центральной нервной системе существуют особые структуры, специализированные на функции торможения и что торможение по своей физико-химической природе противоположно возбуждению. Другие считали, что торможение в центральной нервной cистеме возникает в результате конфликта нескольких возбуждений или же вследствие чрезмерно сильного или длительного возбуждения («перевозбуждения»), т. е. складывается по механизму пессимума Введеского.
Современные электрофизиологические исследования Дж. Экклса, Д. Пурпуры, П. Г. Костюка и др. позволили установить, что в известной мере правы были как те, так и другие следователи, поскольку в центральной нервной системе существует несколько видов торможения, имеющих разную природу и различную локализацию.
- Постсинаптическое торможение
- Пресинаптическое торможение
- Пессимальное торможение в нервных центрах. Торможение деятельности нервной клетки может осуществляться и без участия особых тормозящих структур. В этом случае торможение развивается в возбуждающих синапсах в результате сильной деполяризации постсинаптической мембраны под влиянием слишком частого поступления к ней нервных импульсов. Прообразом такого торможения является пессиум Введенского в нервно-мышечном соединении. К пессимальпому торможению особенно склонны промежуточные нейроны спинного мозга, нейроны ретикулярной формации и некоторые другие клетки, в которых деполяризация постсинаптической мембраны при частом ритмическом раздражении может быть такой интенсивной и стойкой, что в клетке развивается состояние, подобное катодической депрессии Вериго.
- Торможение вслед за возбуждением. Особым видом торможения является торможение, развивающееся в нервной клетке после прекращения ее возбуждения. Оно возникает в том случае, если после окончания вспышки возбуждения в клетке развивается сильная следовая гиперполяризация мембраны. Возбуждающий постсинаптический потенциал в этих условиях оказывается недостаточным для критической деполяризации мембраны, и распространяющееся возбуждение не возникает.
